EL ORIGEN DE LA HOMOQUIRALIDAD BIOLOGICA Y EL NACIMIENTO DE LAS ESTRELLAS.
Por David Serquera Peyró.
Actualmente podríamos dividir las hipótesis que intentan explicar el origen de la vida en dos grandes grupos: Las teorias que preconizan un origen a partir de moléculas orgánicas preformadas en disolución y las que únicamente contemplan un origen autocatalítico progresivo de las biomoléculas. Al primer grupo pertenecen la teoria de la sopa primigenia de Oparin-Haldane soportada por los experimentos de Miller, y la teoría de las arcillas de Bernal en la que tras una unión débil por adsorcion de las biomoléculas a sustancias arcillosas éstas pasarían a fomar parte de la sopa primordial. En frente, la teoría de Günther Wächtershäuser del metabolismo de superficie, en la que se hace necesario un metabolismo autocatalítico sobre una superficie mineral cargada sobre la que se van creando las biomoléculas. Cualquiera de los dos grupos de hipótesis debe dar explicación al origen de la homoquiralidad biológica (suponiendo válido el principio de la inhibición enantiomérica cruzada) permitiendo la teoria de Wachtershauser un origen exclusivamente terrestre para el exceso de L-aminoácidos y D-azúcares, por el contrario, la sopa primigenia admite el origen extraterrestre del exceso unilateral de enantiómeros. En este artículo revisaremos las observaciones científicas que apoyan el origen extraterrestre de la homoquiralidad biológica. Dejaremos para artículos venideros la exposición de los ingeniosos argumentos a favor de una homoquiralidad de origen terraqueo asociada a la estructura de la pirita propuesta por Wachtershauser.
El 28 de Septiembre de 1969, aproximadamente a las 10:45h., los habitantes de una pequeña localidad situada al norte de Melbourne (Australia), llamada Murchison, se vieron sorprendidos por una lluvia de meteoritos que se depositó cubriendo aproximadamente una superficie de cinco millas en forma de pequeños fragmentos de piedra espacial. Se recuperaron más de 100 Kg. siendo el fragmento de mayor tamaño de un peso aproximado de 7 Kg. El meteorito de Murchison pertenece al tipo de los llamados condritas carbonaceas tipoII (CM2) por estar formado fundamentalmente por compuestos del carbono y agua junto con amonio, y óxidos de Azufre y Hierro. La textura se asemeja a algunas rocas volcánicas formadas en la tierra que han sufrido procesos de fragmentación y recimentación y presentan pequeñas formaciones que asemejan fósiles, sin embargo, lo más interesante de este tipo de meteoritos es que se cree que no han sufrido cambios desde su formación en los albores de nuestro sistema solar y tras análisis bioquímicos se ha detectado la presencia de aminoácidos a concentraciones no desdeñables, el hallazgo se hace interesante si pensamos que nosotros estamos formados también en gran parte por aminoácidos, que son las unidades básicas que forman las proteínas y que, éstas, a su vez, son los materiales y las máquinas que crean nuestro orden particular que nos permite existir sobre la Tierra.
El meteorito de Murchison se hace especialmente relevante para el estudio de los orígenes de la vida no sólo por el hallazgo de aminoácidos en su composición, sino, porque además, existe un exceso de L-aminoácidos frente a una menor concentración de D-aminoácidos siendo la primera evidencia natural de la existencia de un posible mecanismo prebiótico de amplificación enantiomérica (1). Pero qué es un L-aminoácido y qué importancia tiene su presencia a altas concentraciones en objetos celestes.
A principios del siglo XVIII se descubre
la posibilidad de polarizar la luz en un plano mediante el llamado prisma de Nicol,
en honor a su descubridor, el físico británico William Nicol, quien creó un mecanismo
formado por espato de Islandia (carbonato cálcico cristalino) mediante el cual
se pudo estudiar el efecto causado por determinadas sustancias sobre un haz de
luz polarizada.
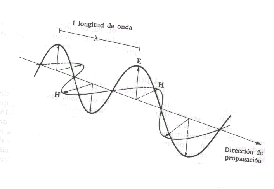
La
luz tiene un comportamiento ondulatorio formado por un campo eléctrico y un campo
magnético perpendiculares entre si, la propagación de estos campos ondulatorios
a través de cualquier medio se realiza con todos los ángulos posibles excepto
si la luz atraviesa determinadas sustancias (materiales polarizadores) que solo
dejan pasar un haz con un determinado plano de ondulación, a este haz de radiación
electromagnética resultante se le llama luz polarizada.
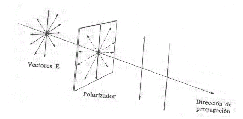
Al atravesar la luz polarizada una sustancia química, sus campos eléctricos y magnéticos interaccionan con las nubes electrónicas de los orbitales moleculares del compuesto lo que induce un cambio en el ángulo del plano de ondulación, se produce una desviación de la luz polarizada que se puede medir con un aparato muy sencillo, llamado polarímetro, cuantificando el ángulo y el sentido de dicha desviación. La desviación de la luz polarizada puede ser en sentido de las agujas del reloj (+), o en sentido contrario a las agujas del reloj (-). De Aquellos compuestos que tienen la capacidad de desviar la luz polarizada en un plano se dice que tienen actividad óptica y son de dos tipos Dextrógiros (+) y levógiros (-).
¿Por qué determinadas sustancias muestran actividad óptica y otras
no? Imaginemos al gran científico francés Louis Pasteur bebiendo una copa de un
excelente Bordeaux mientras se mira en un espejo y tendremos una simpática imagen
de las reflexiones de este gran hombre en torno a uno de sus ingeniosos experimentos
que dio luz a la respuesta definitiva ofrecida posteriormente por Vant Hoff y
Le Bel .
Pasteur conocía que tras el proceso de fermentación del vino en las cubas se depositaban dos ácidos, uno con actividad óptica dextrógira llamado ácido tartárico, y otro sin actividad óptica llamado entonces ácido racémico. Preparó varias sales de dichos ácidos y observó que sus cristales no eran simétricos, eran quirales. La palabra quiral proviene del griego y significa mano. ¿ Alguna vez has visto a un zurdo dando la mano a un diestro? Si miro el reflejo de mi mano que proyecta un espejo veo que ya no es mi mano derecha, es mi mano izquierda. Mis dos manos parecen iguales, tienen cinco dedos, una palma y un dorso semejantes, pero qué pasa si intento superponerlas en la misma orientación espacial , por ejemplo mirando a la vez los dos dorsos. No encajan, son asimétricas, son quirales: tienen la misma forma, están compuestas por los mismos tejidos pero su imagen especular no es superponible. Los cristales del ácido tartárico eran todos semejantes entre si, sin embargo, los cristales del ácido racémico poseían conformaciones diferentes, habían dos tipos de conformación que se relacionaban como imágenes especulares entre sí no superponibles. Pasteur hizo dos grupos, uno con cada conformación, y los sometió por separado al polarímetro encontrando que cada uno de los grupos poseía actividad óptica, un grupo poseía una rotación específica igual a la del ácido tartárico mientras que el otro grupo poseía la misma rotación pero de sentido contrario con lo que Pasteur concluyó que el ácido racémico no era más que una mezcla al 50% de moléculas de ácido tartárico levógiras y dextrógiras con lo que la actividad óptica resultante sería nula. Pasteur fue el primero en descubrir que la actividad óptica está relacionada con la quiralidad, pero ¿de qué forma? Fueron Vant Hoff y le Bel quienes por separado y de manera independiente dieron con la solución. Los compuestos orgánicos están formados por átomos de carbono, el átomo de carbono se puede unir a cuatro átomos más formando un tetraedro, si un átomo de carbono se une a cuatro átomos diferentes se convierte en una estructura geométrica asimétrica en la que sus imágenes especulares (enantiómeros) no son superponibles, así si una sustancia orgánica posee en su estructura al menos un átomo de carbono asimétrico entonces poseerá actividad óptica.
Los aminoácidos poseen un átomo de carbono asimétrico en su estructura por lo que tienen conformaciones especulares diferentes no superponibles que se denominan L-aminoácidos y D-aminoácidos ( por ejemplo hay dos aminoácidos Valina , uno en forma D y otro en forma L). Su actividad óptica depende de su configuración espacial pero no está especificada por ésta; un L-aminoácido puede ser dextrógiro y otro L-aminoácido puede ser levógiro. En la naturaleza todas las proteínas están formadas exclusivamente por L-aminoácidos, no se admite la posibilidad de una proteína formada solo por D-aminoácidos o por una mezcla de D y L-aminoácidos. Esto ocurre también con los azúcares, esta vez en conformación D. A esto se le llama homoquiralidad biológica y es uno de los misterios más apasionantes que nos quedan por desvelar y que con su explicación numerosas cuestiones en torno al origen de la vida serán entendidas.
¿La homoquiralidad biologica es un prerequisito para la vida? ¿Por qué L- aminoácidos y no D? ¿Es producto de una ventaja evolutiva ? ¿Es consecuencia de un mundo prebiótico particular que ha determinado la vida? ¿Son estables las proteínas con una composición heterogenea de enantiómeros? Estas y muchas otras preguntas trataremos de resolver mediante la cuidadosa observación ciéntífica y el ingenio necesario para desvelar la naturaleza.
Según los trabajos de Joyce et. al. (2) en los que se recrea la replicación en etapas tempranas del mundo de RNA, una cadena homoquiral se replica normalmente hasta que se incorpora un enantiómero de distinta conformacíon, produciendose entonces una inhibición enantiomerica cruzada de la síntesis de la cadena homoquiral, de esto se deduce que debió existir algún mecanismo prebiótico que facilitara la incorporación de un solo y determinado enantiómero.
Se han propuesto múltiples mecanismos de selección prebiótica quiral (3). En el laboratorio se han obtenido cantidades importantes de un enantiómero determinado, a partir de mezclas racémicas, mediante la fotolisis asimétrica con luz polarizada circularmente (LPC). El problema es que todos los efectos propuestos son de muy escasa intensidad tal y como se dan en la tierra, esto junto con el descubrimiento del exceso de L-aminoácidos encontrados en el meteorito de Murchison hicieron que Bonner planteara la necesidad de un mecanismo de amplificación situando el origen de la homoquiralidad fuera de la tierra(3).
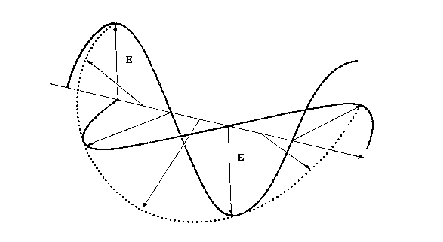
Cuando se superponen dos ondas de luz polarizadas en un plano desfasadas en un cuarto de longitud de onda y con sus vectores E perpendiculares entre sí, se forma la LPC. A medida que se propaga la onda el vector E resultante describe una rotación que forma una curva helicoidal. Si rota en sentido de las agujas del reloj se dice que está polarizada hacia la derecha, si lo hace en sentido contrario se dice que está polarizada a la izquierda.
El mecanismo principal
de generación de un exceso de enatiómeros sería la fotolisis por LPC. Bonner propone
como fuente de esta luz la radiación sincrotónica de púlsares o estrellas de neutrones.
El ejemplo más fácil de hallar es la nebulosa del cangrejo y su púlsar, sin embargo,
para que un aminoácido pueda absorber este tipo de luz ésta debe tener la longitud
de onda del ultravioleta y en este sistema no se ha encontrado LPC UV. Además
si ésta existiese sería de ambos sentidos con lo que la LPC resultante tendria
un efecto neto muy pequeño(4).
Bailey et. al. (5) proponen dos fuentes de LPC distintas. Cuando moléculas esféricas de pequeño tamaño son bombardeadas por un haz de luz polarizada en un plano se produce un fenómeno de difusión (Scattering) por lo que se genera LPC, si las partículas no son esféricas pero están alineadas por un campo magnético como ocurre el la region de formación estelar de la nebulosa de Orión, también se produce por el mismo efecto LPC.
Cuando una perturbación, por ejemplo una explosión de una supernova, actúa sobre estos cúmulos de polvo y gas, se pone en marcha el proceso de formación estelar; la materia se contrae, comienza a rotar y se expulsa en chorros a lo largo del eje de rotación, va siendo atrapada y expulsada quedando una mínima parte libre de atracción formándose los planetesimales. La materia expulsada está alineada por los campos magnéticos en un determinado sentido y la luz incidente de otras estrellas próximas provoca la LPC que incide sobre las biomoléculas provocando su fotolisis asimétrica según la geometría particular del sistema. Durante la fase de bombardeo de la tierra por meteoritos éstos aportarían una ingente cantidad de enantiómeros particulares provocando un exceso homoquiral que determinaría la arquitectura vital de hoy.
En el laboratorio se obtiene un exceso enantiomérico de aminoácidos con 1.8 x 10 elevado a 17 fotones por cm a la -2. Según los cálculos de Bailey et. al. Una estrella A5 en la zona de formacion estelar de Orion aporta 3x10 elevado a nueve fotones por cm a la -2 por segundo a la -1 en un ancho de 30 a 220nm. Una difusión del 0.1% en forma de LPC produciría una irradiación comparable a la de los experimentos en 2000 años.
Quedan sin embargo algunos problemas por resolver, por ejemplo, una vez producido el exceso enantiomérico como disminuiría éste por la progresiva ramecización. Bailey da algunas cifras al respecto en (5). Otro problema importante es la incompatibilidad entre la hipótesis que actualmente explica la formación de aminoácidos en los meteoritos por la sintesis de strecker y la producción del exceso de enantiómeros por LPC, a este respecto Cronin da algunos datos en (1). En frente encontraremos otro camino que nos llevará a buscar el origen de la homoquiralidad en las fumarolas oceánicas sobre una cuna de hierro y azufre, pero eso será en próximos artículos.
BIBLIOGRAFIA:
1. Cronin, Pizzarello. Enantiomeric Excesses in Meteoritic Amino Acids. Science.275.951-55.
2. G.F.Joyce et. Al. Chiral selection in poly(C)-directed synthesis of oligo(G).
Nature. 310. 602-605.
3. W. A. Bonner, Orig. Life. 21, 59 (1991).
4.
Mason. Nature 389, 804(1997).
5. Bailey et. Al. Circular Polarization in
Star-formation regions: Implications for biomolecular homochirality. Science.281.672-674.
6. W. A. Bonner. Chirality Amplification-The accumulation principle revisited.
Origins of life.29.615-623,1999.
7. T. P. Ray. Los primeros días de la vida
de una estrella. Investigación y ciencia. Octubre 2000.
8. R. L: Dickman Los
glóbulos de Bok. Investigación y ciencia.
9. Quimica Fisica. Díaz Peña. Ed.
Alhambra.
10. Química Orgánica. Hart. Ed. Mc Graw Hill.
11. Técnicas
de Bioquímica y Biología molecular. Freifelder. Ed Reverté.
12. Orígenes
de la evolución biológica. Juli G. Peretó. Ed. Eudema.